
L'activation de l'ADN : un déplacement entre deux territoires nucléaires ?
L'ADN est souvent décrite comme de longs brins d'ADN et visualisé comme une sorte de X ressemblant à 2 merguez attachées en leur centre, mais des recherches récentes révèlent une organisation tridimensionnelle insoupçonnée dans le noyau interphasique qui pourrait être liée à l'activation des gènes. Dans ce nouveau cadre théorique, l'équipe du professeur Duboule de l'Université de Genève a publié tout récemment une recherche pourrait faire date : en montrant que l'activation successive des gènes architectes Hox dans l'ordre de leur position sur le chromosome correspond au passage successif d'un compartiment négatif "off" à un compartiment positif "on" à l'intérieur du noyau.
Il pourrait s'agir d'un mécanismes général pour expliquer au moins certaines régulations épigénétiques.
Avec d'autres travaux sur le positionnement de l'ADN dans le noyau, il semble se dégager un modèle très élégant et simple de (certaines?) activations du génome : Simplement passer d'un compartiment à l'autre placerait l'ADN dans un environnement biochimique différent qui l'activerait ou le désactiverait. Les modifications de la chromatine comme la méthylation seraient un effet de cet environnement ? Il est prématuré d'être trop affirmatif, mais il me semble pressentir un de ces rares moments où les savoirs très éclatés que produit la recherche convergent d'un coup vers un modèle relativement simple.
Un ADN linéaire ou replié en "merguez" ?
L'ADN est souvent décrit comme un long brin d'ADN – ou 46 long brins – et la biologie actuelle qui aborde de nombreux problèmes par l'étude des séquences fait référence à une structure linéaire. Selon Prof Bruno Strasser, de l'UniGE cette vision, formulée il y a plus de cinquante ans par Francis Crick comme un "dogme central", a été au coeur de la recherche en biologie moléculaire depuis lors (Strasser, B. J. 2006). Cette perspective, qui réduit l'ADN a sa séquences, a permis l'émergence d'une biologie de l'information plutôt que de la forme. Mais, comme le montrent certains travaux récents, cette vision est probablement trop simpliste.C'est que l'information pure est peu visuelle et l'enseignement de la biologie a ses traditions...
En effet on représente en général les chromosomes par une figure en X comme deux "merguez" liées au centre. En principe, on explique aux élèves que la plupart du temps la cellule a ses chromosomes non condensés, en "vrac" dans le noyau sous forme de "chromatine". Et que cette représentation en X correspond à la métaphase, un moment très bref de la division cellulaire.
Mais cette image en X est si prégnante qu'elle s'impose partout comme représentation du chromosome, même pour faire référence à d'autres phases du cycle cellulaire, qui sont la très grande majorité du temps de la cellule. C'est une des représentations du matériel génétique les plus utilisées.

Fig 1 : La forme en "2-merguez" s'est imposée comme représentation stéréotypée du chromosome ou de l'ADN voire du matériel génétique. [ img ]Source :inconnue désolé. merci de me signaler si vous trouvez.
Certains schémas cherchent à articuler ces deux représentations : linéaire et "en 2-merguez".
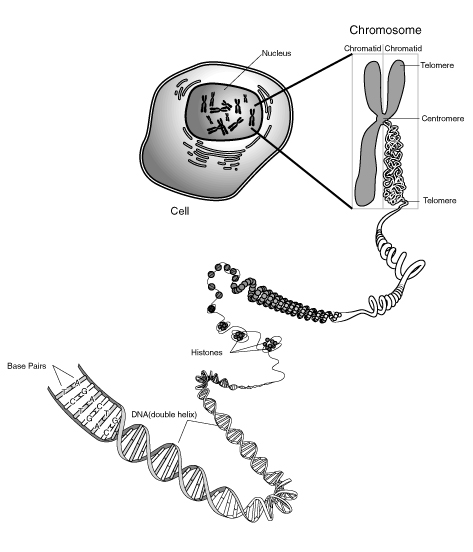
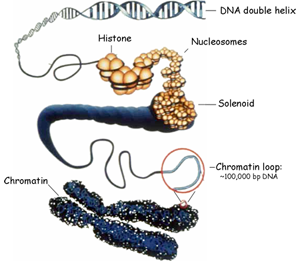
Fig 2 : Certains schémas cherchent à articuler ces deux représentations : linéaire et "en 2-merguez". Gauche [ img ]Source : wikipedia Droite [ img ] : Source ici
Evidemment dessiner l'ADN durant l'interphase est difficile et s'apparente plus aux gribouillis d'un enfant qu'à un schéma scolaire facile à retenir... certains schémas le tentent cependant et montrent bien les difficultés que cela pose.

Fig 3 : Certains schémas tentent de représenter l'ADN durant l'interphase. Au risque de suggérer que l'ADN sort du noyau comme ici. [ img ]Source : http://mcb.illinois.edu/faculty/profile/cmizzen
Une des raisons de cette difficulté est qu'on en savait pas vraiment comment – ou même si – l'ADN était organisé dans le noyau, entre autres parce qu'ils n'étaient pas discernables au microscope. Sauf durant la mitose et la méiose, et tout particulièrement bien à la métaphase... Ce qui explique probablement le succès des "merguez"
L'ADN est la plupart du temps en vrac dans le noyau ?
Pendant longtemps on a ignoré tout de l'organisation des chromosomes dans le noyau qui ne se divise pas. (Meaburn, K. J., & Misteli, T. 2007).On sait depuis quelques années que l'ADN s'organise dans le noyau de manière complexe mais plutôt groupée par chromosome. Une News And Views de Nature fait le point sur les "Chromosome territories" (Meaburn, K. J., & Misteli, T. 2007):
Fig 4 : Déjà en 2007 on savait que les chromosomes interphasique ne sont pas en vrac dans le noyau. a) par fluorescence on a coloré le chromosome 12 (rouge), chromosome 14 (vert) and chromosome 15 (bleu). b) chaque chromosome est coloré différemment, c) Les territoires des chromosomes ne sont pas compacts. [ img ] Source : Meaburn, K. J., & Misteli, T. (2007) :

Des articles de "Nature" pour les élèves ? vous n'y pensez pas !
Ces infos sont reprises et mises à jour dans un article spécialement préparé pour l'éducation proposé dans le site éducation de Nature Scitable . Misteli, T. (2008) Chromosome territories: The arrangement of chromosomes in the nucleus . Nature Education 1( 1 )Fig 5 : Visualization of chromosome territories by fluorescence in situ hybridization . B) La position des chromosomes 12, 14 et 15 est indiquée. [ img ]Source : Meaburn, K. J., & Misteli, T. (2007)

Evidemment c'est en anglais ... mais c'est une excellente ressource – gratuite – de documents bien plus authentiques.
L'efficacité pour motiver les élèves de documents extra-scolaires et authentiques à été établie notamment par le prof. (Müller, A.,et al. (2010) de l'UniGE.
On peut d'ailleurs noter que juste après le secondaire ils vont souvent à l'université (!) où la lecture de l'anglais est une exigence en sciences...
Une structure 3-D liée à l'expression des gènes ?
Selon Baker, M. (2011) le prochain horizon en génomique est l'espace : l'organisation tridimensionnelle des chromosomes dans le noyau. Ce n'est pas simplement 2m d'ADN tassés au hasard dans une sphère de 10 micromètres, et les structures varient selon les types de cellules et semblent jouer un rôle encore mal compris dans l'expression des gènes.
Fig 6 : Une structure magnifiquement illustrée dans d'un fragment de 500-kilobase riche en gènes sur le chromosome 16 humain.[
img
]Source :Baker, M. (2011) J. DEKKER/UNIV. MASSACHUSETTS/D. BAU/M. A. MARTI-RENOM/PRINCE FELIPE RES. CENTRE.
On sait depuis un certain temps que les interactions qui régulent l'expression de l'ADN se font parfois avec des séquences apparemment à des distances importantes. Tant qu'on a pensé l'ADN comme un simple séquence on a eu de la peine à conceptualiser comment les séquences aussi éloignées pouvaient participer à la régulation des gènes. D'imaginer qu'elles se replient a permis de comprendre certaines interactions. L'équipe du Prof Strubin de l'UniGe avait participé à comprendre l'importance potentielle de ces interactions en montrant que si on aide artificiellement la machine de transcription à se fixer au début du gène, on augmentes la transcription du gène en question. (Klages, N., & Strubin, M., 1995).
-
5 manières dont la régulation de l'expression des gènes peut se faire :
Figure 7.49
( Alberts B., et al., 2002)
On-line
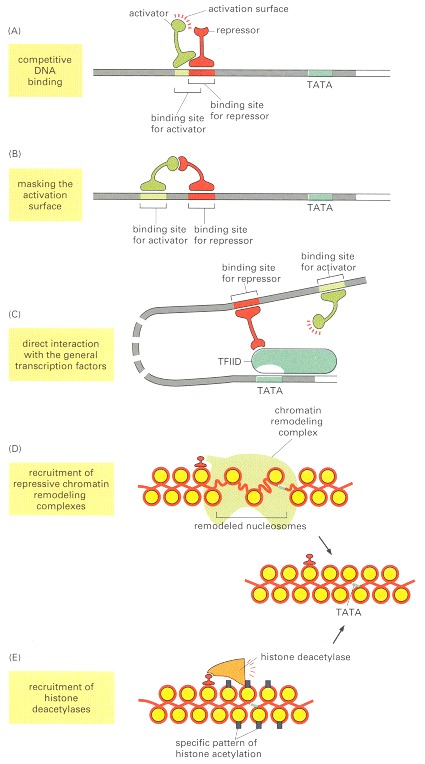
Fig 7: Tant qu'on a pensé l'ADN comme un simple séquence on a eu de la peine à conceptualiser comment les séquences aussi éloignées pouvaient participer à la régulation des gènes.[ img ]Source : Alberts
On sait aussi que la méthylation inactive des gènes (Alberts B., et al., 2002 ici) de manière durable et le domaine en grand développement de l'épigénétique
On commence a connaitre le mécanisme d'inactivation du Chromosome X décrit dans un très bon review des liens entre l'organisation nucléaire et la régulation génique (Fraser, P., & Bickmore, W. (2007). doi: 10.1038/nature05916 ) Notamment la manière dont différentes parties du même chromosome doivent rester actives pour inactiver le reste dans le cas du Chromosome X.

Fig 8: Les évènements de réorganisation nucléaire lors de l'inactivation du chromosome X sont décrits ici .[ img ]Source : Fraser, P., & Bickmore , W. (2007)
Un compartiment actif et un compartiment inactif ?
La conformation globale du génome dans le noyau a été établie récemment par Lieberman-Aiden, E., et al. (2009).Ils ont utilisé une technique HiC qui relie des ADN proches avec des sortes de menottes (formol) , relie ces fragments "pris sur le fait" d'être ensemble puis séquence ces fragments pour identifier quelles séquences étaient proches.
Plus précisément, elle consiste à cross-linker les brins d'ADN qui se trouvent proches avec du formaldéhyde, puis à couper en fragments avec une enzyme de restriction ces ADN attachés, puis à relier les extrémités libres des deux fragments d'ADN et les marquer avec de la biotine pour ensuite séquencer ces fragments et identifier leurs positions dans le génome avec des outils bioinformatiques. La technique a depuis été améliorée et est nommée 3C ( chromosome conformation capture) puis 4C quand elle est associée à du séquençage a large échelle ou des microarray.

Fig 9 La technique Hi-C consiste à cross-linker les brins d'ADN qui se trouvent proches avec du formaldéhyde, puis à couper en fragments avec une enzyme de restriction ces ADN attachés, puis à relier les extrémités libres des deux fragments d'ADN et les marquer avec de la biotine pour ensuite séquencer ces fragments et identifier leurs positions dans le génome avec des outils bioinformatiques. . [ img ]Source : Lieberman-Aiden, E., et al. (2009)
Leur étude montre qu'il y a deux compartiments dans le noyau : des zones où la chromatine est ouverte et d'autres où elle est fermée.
" Our results demonstrate that open and closed chromatin domains throughout the genome occupy different spatial compartments in the nucleus. "Sans tenter ici de faire un review complet de l'épigénétique (CF ici ) on peut relever qu'une séparation spatiale permet d'imaginer des mécanisme très simple pour expliquer l'activation / la désactivation de parties du génome.
Une structure fractale rapproche tout le génome et permet de passer de l'un à l'autre
Leur étude montre aussi que s'établissent des contacts aussi bien entre des zones proches dans l'ADN qu'entre des zones très lointaines : ils la qualifient de globule fractal parce les rapprochements se répètent à des niveaux d'organisation différents ce qui explique qu'il y a autant de chances de rencontre entre des séquences proches que très lointaines. Ce serait pour permettre l'activation / désactivation des gènes en passant d'un compartiment à l'autre."At the megabase scale, the chromatin conformation is consistent with a fractal globule, a knot-free, polymer conformation that enables maximally dense packing while preserving the ability to easily fold and unfold any genomic locus. The fractal globule is distinct from the more commonly used globular equilibrium model." Lieberman-Aiden, E., et al. (2009)

Fig 10 D es contacts s'établissent aussi bien entre des zones proches dans l'ADN qu'entre des zones très lointaines : ils la qualifient de globule fractal (C en bas) parce les rapprochements se répètent à des niveaux d'organisation différents (D) ce qui explique qu'il y a autant de chances de rencontre entre des séquences proches que très lointaines. [ img ] Source : Lieberman-Aiden, E., et al. (2009) .
Notre société gagnerait à être fractale ?
La société aurait peut-être bien besoin d'une structure fractale qui rapproche les différentes groupes si éloignés qui la composent et rend facile le passage d'un état d'activation à l'autre...Et peut-être aussi une éducation qui respecte les différences mais donne à chacun sa chance de quitter état silencieux pour commencer à s'exprimer quelle que soit sa position sociale ?
Les gènes Hox s'activent dans l'ordre en passant du compartiment "off" au compartiment "on"
Les gènes architectes Hox spécifient les structures le long de l'axe antéro-postérieur du corps. Ils s'expriment dans un ordre qui correspond aussi bien à leur position sur les chromosomes qu'à l'ordre des structures corporelles. . Ce qui détermine cette activation successive n'était pas simple à expliquer...-
Cf. publication
Bio-Tremplins du 5 avril 2011
 pour approfondir.
[img]
pour approfondir.
[img]
Dans la revue Science du 14 octobre 2011 L'équipe du professeur Duboule de l'Université de Genève publie une recherche qui pourrait faire date : en montrant par la technique (4C similaire à Hi-C décrite plus haut) le passage successif des gènes architectes Hox d'un compartiment négatif "off" à un compartiment positif "on" dans le noyau on suggère un modèle très élégant et simple de leur activation successive dans l'ordre de leur position.
Simplement passer d'un compartiment à l'autre situe le brin d'ADN et ses histones dans un environnement biochimique différent qui les activerait ou les désactiverait.
This bimodal configuration parallels the distribution of distinct chromatin marks, suggesting the existence of a link between the presence of chromatin domains and the formation of 3D chromosomal structures. This model for Hox gene activation would ensure the proper sequence in the transcriptional activation of Hox genes within each gene cluster. Noordermeer, D., Leleu, M., Splinter, E., Rougemont, J., De Laat, W., & Duboule, D. (2011).

Fig 11 : Cette étude révèle passage successif des gènes Hox d'un compartiment négatif "off" (rouge9 à un compartiment positif "on" (Bleu) dans le noyau, de manière correspondante à leur activation. [ img ]Source : Noordermeer, D., Leleu, M., Splinter, E., Rougemont, J., De Laat, W., & Duboule, D. (2011). :
"This work suggests that the colinear activation of Hox genes involves a stepwise transition of each gene from a negative to a positive compartment, which display different biochemical properties and thus results in a physical separation of their regulatory modalities. " Noordermeer, D., Leleu, M., Splinter, E., Rougemont, J., De Laat, W., & Duboule, D. (2011)Ainsi de considérer le problème en 3-D a probablement permis à certains d'imaginer la nouvelle technique qui a permis à d'autres de faire de nouvelles expériences qui conduisent à revoir les modèles explicatifs...
Un nouveau modèle des phénomènes mène à de nouvelles expériences qui construisent et affinent le modèle. C'est le processus de modélisation. Dans les moments où ils se construisent, on perçoit bien les caractéristiques du modèle (ils sont hypothétiques, ils sont modifiables, ils sont pertinents pour certains problèmes dans certains contextes. (Martinand, J. L., 1996)). Une fois bien établis on finit souvent par les présenter et les utiliser comme "vrais", définitifs, inébranlables, absolus.
Puis on s'étonne que les élèves ne distinguent pas science et dogme.
Un moment historique ?
Il semble donc qu'un changement de cadre conduise à un nouveau modèle : une vision de l'ADN en 3-D dans le noyau a permis d'aller beaucoup plus loin dans la compréhension, par une sorte de retour ironique de l'histoire des idées.En effet c'est l'idée de simple linéarité de Crick et de flux d'information qui avait permis de voir autrement les données et de comprendre sous l'angle d'une biologie de l'information, génomique et bioinformatique notamment.
Et voilà qu'au coeur de l'information les mécanismes de régulation se comprennent bien mieux en examinant la conformation tridimensionnelle du génome dans le noyau. Ce que l'analyse des séquences ne montre pas, ou pas d'une manière que nous comprenons.
Si cela se confirme nous serions en train de vivre un de ces rares moments où les savoirs très éclatés que produit la recherche convergent d'un coup vers un modèle relativement simple. Qui pourrait assez simplement être présenté aux élèves. C'est en tous cas intéressant de montrer que la science bouge !
Evidemment cette réponse suggère d'autres questions : qu'est-ce qui déplace l'ADN d'un compartiment vers l'autre ? Qu'est-ce qui détermine ces territoires ? des gènes ? etc.
Magnifiques questions pour les jeunes chercheurs parmi nos élèves.
"Scitable is a free science library and personal learning tool brought to you by Nature Publishing Group, the world's leading publisher of science. Scitable currently concentrates on genetics and cell biology , which include the topics of evolution, gene expression, and the rich complexity of cellular processes shared by living organisms. Scitable also offers resources for the budding scientist, with advice about effective science communication and career paths ." |
Le site éducation de Nature
Scitable

Fig 12 : Scitable est le site éducation de Nature . [ img ]Source : Nature Scitable
Sources
-
Alberts B, Johnson A, Lewis J, et al. (2002) Molecular Biology of the Cell . 4th edition.New York: Garland Science; .
- Baker, M. (2011). Genomics: Genomes in three dimensions. Nature, 470(7333), 289-294. doi: 10.1038/470289a | intranet.pdf
- Crick, F. (1970). Central dogma of molecular biology. Nature, 227(5258), 561-563.
-
Fraser, P., & Bickmore, W. (2007). Nuclear organization of the genome and the potential for gene regulation. Nature, 447(7143), 413-417. doi:
10.1038/nature05916
- Lieberman-Aiden, E., et al. (2009). Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome. Science, 326(5950), 289 -293. doi: 10.1126/science.1181369
- Martinand, J. L. (1996). Introduction à la modélisation. Paper presented at the Actes du séminaire de didactique des disciplines technologiques. extraits intranet.pdf
-
Meaburn, K. J., & Misteli, T. (2007). Cell biology: Chromosome territories. Nature, 445(7126), 379-381. doi:
10.1038/445379a
- Misteli, T. (2008) Chromosome territories: The arrangement of chromosomes in the nucleus . Nature Education 1( 1 )
-
Müller, A., Kuhn, J., Müller, W., & Vogt, P. (2010). `Modified Anchored Instruction´ im Naturwissenschaftlichen Unterricht: Ein Interventions- und Forschungsprogramm. In D. Höttecke (Ed.), Entwicklung naturwissenschaftlichen Denkens zwischen Phänomen und Systematik, GDCP-Tagungsband 2009. Münster: LIT.
- Sexton, T., Schober, H., Fraser, P., & Gasser, S. M. (2007). Gene regulation through nuclear organization. Nat Struct Mol Biol, 14(11), 1049-1055. doi: 10.1038/nsmb1324
- Noordermeer, D., Leleu, M., Splinter, E., Rougemont, J., De Laat, W., & Duboule, D. (2011). The Dynamic Architecture of Hox Gene Clusters. Science, 334(6053), 222 -225. doi: 10.1126/science.1207194
- Strasser, B. J. (2006). A world in one dimension: Linus Pauling, Francis Crick and the central dogma of molecular biology. History and Philosophy of the Life Sciences, 28, 491-512. article.pdf
-
Klages, N., & Strubin, M. (1995). Stimulation of RNA polymerase II transcription initiation by recruitment of TBP in vivo. Nature, 374(6525), 822-823. doi:
10.1038/374822a0
- Zimmer, C., & Fabre, E. (2011). Principles of chromosomal organization: lessons from yeast. The Journal of Cell Biology , 192 (5), 723 -733. doi:10.1083/jcb.201010058





Aucun commentaire:
Enregistrer un commentaire